
琥珀酸脱氢酶抑制剂(succinate dehydrogenase inhibitor,SDHI)类杀菌剂是一类新型杀菌剂,始于1965年,首例是由Uniroyal公司研发并申请专利,可用于防治单胞锈菌Uromyces vignae引起的菜豆锈病和大麦坚黑粉菌Ustilago hordei引起的大麦黑穗病。SDHI类杀菌剂因具有低毒、高效、安全、广谱等特点而被广泛用于农业中。例如,2022年全球杀菌剂市场价值达191.7亿美元,其中三唑类占比最大,其次为甲氧基丙烯酸酯类和SDHI类,2019年这3类杀菌剂的全球销售额依次为43.67亿、32.12亿、23.11亿美元。而从增长率来看,除生物杀菌剂外,SDHI类杀菌剂的同比增长率及2014—2019年的复合年增长率均最高,分别为5.3%和8.5%。
SDHI类杀菌剂作用位点单一,主要通过作用于蛋白复合体Ⅱ影响病原菌的呼吸链电子传递系统,阻碍其能量代谢,抑制其生长,导致其死亡。杀菌剂抗性行动委员会(Fungicide Resistance Action Committee,FRAC)已将SDHI类杀菌剂归为中等抗性风险杀菌剂,长期、频繁的使用该类杀菌剂会导致病原菌产生抗药性,导致蔬菜灰霉病、小麦壳针孢叶枯病、苹果黑星病、马铃薯早疫病、百合叶枯病和油菜菌核病等病害再猖獗;在中国,小麦赤霉病菌Fusarium graminearum、番茄灰霉病菌Botrytis cinerea、桃腐烂病菌Alternaria alternata和黄瓜靶斑病菌Corynespora cassiicola等病原菌已对SDHI类杀菌剂产生抗性。抗药性的发展可能降低SDHI类杀菌剂对病害的控制,这给未来SDHI类杀菌剂的结构设计和抗性治理带来挑战。因此,该文从发展脉络、生物活性、环境行为、生态毒理等方面对SDHI类杀菌剂进行全面综述,以期为该类杀菌剂的研究和应用提供参考。
1 SDHI类杀菌剂的发展脉络
在过去的50年里,SDHI类杀菌剂凭借其独特作用机制实现了跨越式发展,先后完成从保护性向内吸性、从窄谱抑菌向广谱杀菌的两大历史性转变,成为杀菌剂领域的重要研究与应用方向。依据FARC分类体系,目前已有25种SDHI类杀菌剂,根据化学结构特征这25种SDHI类杀菌剂被归为12个类别,这12个类别是现代农业病害绿色防控体系中的核心药剂品类。
1.1 第1代SDHI类杀菌剂
1966年,1,4-氧硫杂环己二烯酰胺类衍生物被Schmeling发布,并用于化学防治小麦叶面黑穗病、锈病等。1969年美国好利来路公司成功研发出了全球首个SDHI类杀菌剂萎锈灵;1970年Mathere指出萎锈灵的作用靶标为琥珀酸脱氢酶(succinate dehydrogenase,SDH),其对担子菌引起的病害等有良好的防治效果;萎锈灵还可用于防治多种病害,如通过种子处理来抑制水稻黑穗病,叶面喷雾来预防和治理禾谷锈病等。随着萎锈灵的广泛使用,其许多弊端暴露出来,如易被氧化和光降解等。为提高萎锈灵的杀菌效果并降低其对植物的毒性,1975年美国研制出了萎锈灵的衍生物氧化萎锈灵,该杀菌剂可用于园艺作物。从此,各大农化公司纷纷投身于SDHI类杀菌剂的研制。1981年,日本公司研制出了灭锈胺,该杀菌剂能有效防治多种作物的主要病害,而且具有持效期长、不易产生药害、耐雨水冲刷等优点;例如,20%灭锈胺对水稻纹枯病有良好的防治效果。1974年壳牌公司研制出了甲呋酰胺。1986年,日本农药公司研制出了氟酰胺。同年,巴斯夫研发出了麦锈灵。这6种SDHI类杀菌剂防治范围窄,因此并未得到广泛使用。
1.2 第2代SDHI类杀菌剂
为解决杀菌剂防治范围窄的问题,于是第2代SDHI类杀菌剂问世。SDHI类杀菌剂的研究主要集中在结构中,尤其是关于酸部分和胺部分,而关于酰胺键部分的研究相对较少。第2代SDHI类杀菌剂通过替换取代基研制出了新化合物,这推动了SDHI类杀菌剂的更新。1996年日本住友化学公司发现氟吡菌酰胺是SDHI类杀菌剂发展的关键节点,将羧酸部分引入到SDHI类杀菌剂的吡唑环中。另一个靶向SDHI类杀菌剂为孟山都公司于1997年成功开发的噻呋酰胺,可采取叶面喷施、种子包衣和底肥撒施这3个施药方式来防治田间水稻纹枯病。例如,氟酰胺和噻呋酰胺能有效防控水稻纹枯病的发病率,降低水稻纹枯病的发病程度。第2代SDHI类杀菌剂的研发在结构方面取得了巨大进步,作用范围比第1代更广泛,为第3代SDHI类杀菌剂的开发奠定了基础。
1.3 第3代SDHI类杀菌剂
2003年巴斯夫研发出首例第3代SDHI类杀菌剂啶酰菌胺,其具有广谱的杀菌性,主要用于防治白粉病、灰霉病、叶斑病等,在市场上的认可度较高。该类杀菌剂结构中酸部分的变化主要是含氮杂环的衍生,如吡唑、吡啶和噻唑等的优化;胺部分的变化主要是含氮/氧原子的引入。吡唑基取代吡啶基后的化合物广谱性强且高效。因此日本三井株式会社、先正达公司、拜耳公司、日本农药株式会社等各大农药公司纷纷研发,此时SDHI类杀菌剂的发展达到黄金时期。例如,2009年日本三井株式会社研制出了吡噻菌胺,该杀菌剂的官能团中含有吡唑和酰胺,对白粉病、枯萎病以及灰霉病等病害有很好的防治效果;2011年、2013年、2016年先正达公司先后研制出了吡唑萘菌胺、氟唑环菌胺、苯并烯氟菌唑和氟唑菌酰羟胺,在SDHI类杀菌剂方面取得了显著成绩,实现了长期稳定的发展。吡唑萘菌胺是叶面杀菌剂,可用于小麦、杧果树和花椒树等作物病害防治;氟唑环菌胺是种子处理杀菌剂,可用于谷物、油菜、马铃薯等作物病害防治;苯并烯氟菌唑采用叶面喷雾或土壤处理来防治燕麦、小麦和蓝莓等作物病害。2010年和2011年拜耳公司先后研制出了氯氟联苯吡菌胺和氟唑菌苯胺,2020年研制出了异丙氟吡菌胺。2015年日本石原产业株式会社研制出了异丙噻菌胺。2018年日本农药株式会社研制出了联苯吡嗪氟胺。
总的来说,以萎锈灵等为代表的第1代SDHI类杀菌剂虽有一定的防治效果,但存在易氧化、光降解等弊端,且防治范围较窄。第2代SDHI类杀菌剂通过替换取代基研制出了氟吡菌酰胺和噻呋酰胺等新化合物,显著拓宽了SDHI类杀菌剂的作用范围,为后续发展奠定基础。以啶酰菌胺为开端的第3代SDHI类杀菌剂具有广谱的杀菌性,这得益于其结构优化和众多农化公司的持续研发,开发出了吡噻菌胺、吡唑萘菌胺、氟唑环菌胺等新品种,这推动了SDHI类杀菌剂的广泛应用和持续发展。从早期的萎锈灵到如今的广谱杀菌剂啶酰菌胺等,SDHI类杀菌剂在防治多种作物病害方面发挥了重要作用,同时因其独特的作用机制和良好的性能,普遍认为在未来将成为全球杀菌剂市场中的重要部分,有望继续为农业生产保驾护航。
2 SDHI类杀菌剂的生物活性
生物活性是量化SDHI类杀菌剂农药防效的关键指标。SDHI类杀菌剂的作用机制主要涉及对细胞呼吸链中SDH的抑制。SDH是细胞内的关键酶,又称线粒体呼吸链复合体Ⅱ,又称琥珀酸-辅酶Q还原酶,由黄素蛋白、铁硫蛋白以及两个跨膜蛋白4个功能亚基组成。SDH广泛存在于动物、植物、微生物和细胞的线粒体中,催化从琥珀酸氧化到延胡索酸和从泛醌(辅酶Q)还原到泛醇的偶联反应。SDHI类杀菌剂通过抑制SDH来削弱微生物的能量生产能力,这种抑制作用不仅阻碍三羧酸循环的进行,还切断病原菌的能量供应,从而导致其代谢紊乱,进而导致细胞失去正常的代谢和能量平衡,最终导致微生物的死亡。
本文从菌丝生长抑制率、细胞膜通透性、细胞外草酸和多糖含量、SDH活性等方面对SDHI类杀菌剂生物活性差异进行研究。SDHI类杀菌剂共有12种化学结构,分别为苯基环丁基吡啶酰胺类、N-环丙基-N-苄基-吡唑-4-酰胺类、噻吩酰胺类、氧硫杂环己二烯酰胺类、苯基苯甲酰胺类、呋喃酰胺类、吡唑-4-酰胺类、噻唑酰胺类、吡啶酰胺类、吡啶乙基苯甲酰胺、苯基氧代乙基噻吩酰胺类以及吡嗪酰胺类。不同SDHI类杀菌剂对不同菌种的有效中浓度EC50差异巨大(表1)。(1)氧硫杂环己二烯酰胺类SDHI类杀菌剂包括萎锈灵和氧化萎锈灵。李敏等在河南省花生主产区采用菌丝生长速率抑制法测定萎锈灵对28株花生白绢病病原菌齐整小核菌Sclerotium rolfsii菌株的生物活性良好,EC50为0.183 mg/L。(2)苯基苯甲酰胺类SDHI类杀菌剂包括灭锈胺、麦锈灵以及氟酰胺;噻唑酰胺类SDHI类杀菌剂仅有噻呋酰胺。史晓晶等采用菌丝生长速率法探究引起玉米纹枯病的立枯丝核菌Rhizoctonia solani融合群AG-5对氟酰胺、噻呋酰胺和戊菌隆的敏感性,结果表明,氟酰胺对立枯丝核菌融合群AG-5有较好的抑制作用,噻呋酰胺对玉蜀黍丝核菌R.zeae融合群WAG-Z有较高的抑制效果,EC50分别为0.168 mg/L和0.677 mg/L。史晓晶等采用菌丝生长速率法测得噻呋酰胺对禾谷丝核菌R.cerealis AG-DI亚群的EC50范围为0.067~0.142 mg/L。(3)呋喃酰胺类SDHI类杀菌剂仅有甲呋酰胺,关于其研究较少。(4)吡唑-4-酰胺类SDHI类杀菌剂包括氟苯醚酰胺和苯并烯氟菌唑,对多种病原菌均具有较高的抑制效果。例如张俊甜采用离体叶断法测定氟苯醚酰胺对173株小麦条锈菌Puccinia striiformis f.sp.tritici分离菌株的EC50为0.010~0.440 mg/L,平均EC50为0.120 mg/L,结果表明,氟苯醚酰胺对小麦条锈菌有良好的抑制效果,可作为优选杀菌剂。王爱萍等采用菌丝生长速率法测定苯并烯氟菌唑对尖孢炭疽菌Colletotrichum acutatum的菌丝生长的EC50为0.073~3.398 mg/L。(5)吡啶酰胺类SDHI类杀菌剂仅有啶酰菌胺。啶酰菌胺性能好,不仅具有抑制孢子萌发、广谱高效和耐冲刷等优点,而且有较好的预防和治疗效果。例如,汪汉成等采用生物测定法探讨8种杀菌剂对烟草叶斑病菌Didymella segeticola的生物活性,其中啶酰菌胺的抑菌活性最强,代森锰锌的抑菌活性最弱;25 mg/L啶酰菌胺和25 mg/L氟硅唑的防治效果最佳,均大于80.00%,代森锰锌最差,在100 mg/L浓度下其防效仅为63.31%。方安菲等采用菌丝生长速率法测定啶酰菌胺对稻曲病菌Ustilaginoidea virens的EC50介于29.239~85.779 mg/L之间。(6)噻吩酰胺类SDHI类杀菌剂和N-甲氧基-N-苯乙基-吡唑-4-酰胺类化合物都仅有1个,分别为异丙噻菌胺和氟唑菌酰羟胺。刘永春和祁之秋采用离体叶片法和田间药效试验测得氟唑菌酰羟胺、异丙噻菌胺、吡噻菌胺、啶酰菌胺对大樱桃灰霉病菌有良好的抑制效果,EC50介于0.810~7.140 mg/L之间,异丙噻菌胺和氟唑菌酰羟胺对该病害的田间防治效果均大于70.00%,优于其他供试药剂;因此在农业生产中可使用异丙噻菌胺、氟唑菌酰羟胺来防治大樱桃灰霉病。(7)吡啶-乙基-苯甲酰胺类SDHI类杀菌剂仅有氟吡菌酰胺。华学文等探究了9种杀菌剂对苹果炭蛆病菌Colletotrichum aenigma的抑制效果,其中氟吡菌酰胺对苹果炭蛆病菌的抑制率为88.3%,可用于防治苹果炭蛆病。(8)吡嗪-酰胺类、苯基环丁基吡啶酰胺类和N-环丙基-N-苄基-吡唑-4-酰胺类SDHI类杀菌剂均只有1个,分别为联苯吡嗪菌胺、三氟吡啶胺以及异丙氟吡菌胺。例如,侯毅平等测定采自江苏省的105株油菜菌核病菌Sclerotinia sclerotiorum菌株对联苯吡嗪菌胺的EC50介于2.298~0.082 mg/L之间,表明联苯吡嗪菌胺对油菜菌核病的防治效果较好,可以预防油菜菌核病的发生;乔广行等测定三氟吡啶胺、氟唑菌酰胺、氟唑菌酰羟胺和氟啶胺这4种杀菌剂对牡丹黑斑病病原菌交链格孢菌Alternaria alternata的抑制作用,其中,三氟吡啶胺对交链格孢菌菌丝生长的抑制效果最好,EC50为0.270 mg/L,氟唑菌酰胺对交链格孢菌孢子萌发的抑制作用最小,EC50为0.003 mg/L。目前,关于异丙氟吡菌胺的研究主要集中在对水生生物的毒性方向,暂无生物活性方面的研究。
表1 SDHI类杀菌剂代表性化学结构的生物活性
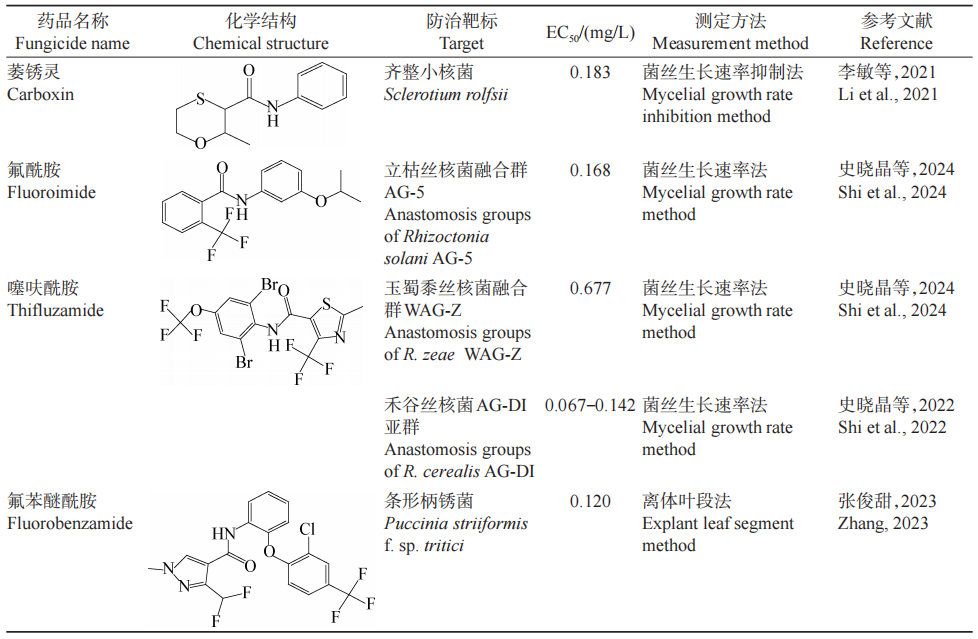
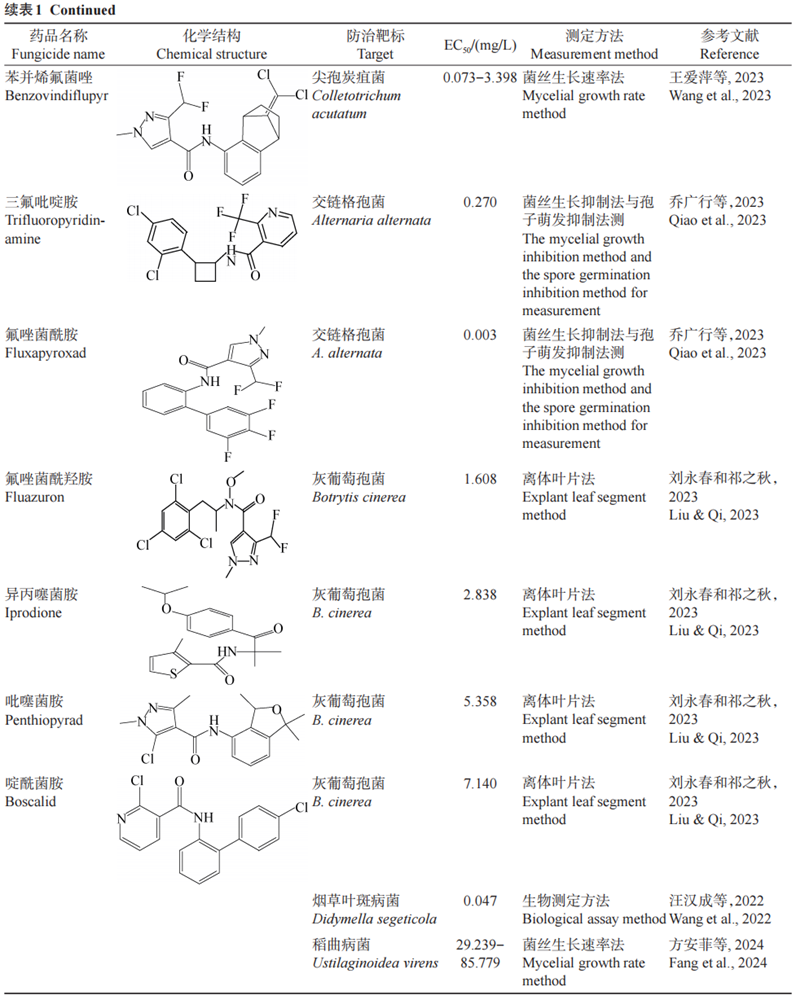
综上所述,SDHI类杀菌剂对不同病菌表现出多样化的生物活性,这与SDHI类杀菌剂的化学结构密切相关。萎锈灵对齐整小核菌、氟酰胺对立枯丝核菌、氟苯醚酰胺对小麦条锈菌等均有较好的抑制作用。此外,啶酰菌胺、氟唑菌酰羟胺等对烟草叶斑病菌和大樱桃灰霉菌的抑制作用较好。然而,关于部分SDHI类杀菌剂如甲呋酰胺和异丙氟吡菌胺的研究相对较少,其生物活性有待进一步挖掘。未来SDHI类杀菌剂的研发应聚焦于抗药性管理方面,开发新型化合物与复配剂以延缓抗药性发展;注重环境友好型配方的创新,降低毒性与残留;开发精准施药技术,如数字化农业与决策支持系统,提升施药效率与精准度。此外,将SDHI类杀菌剂拓展应用至新兴市场与非传统农业领域,将基础与应用研究相结合,加强多学科合作,如利用人工智能辅助新农药创制,推动SDHI类杀菌剂高效、环保的可持续发展。
3 SDHI类杀菌剂的生态毒理
SDHI类杀菌剂对非靶标生物影响的测定通常包括急性毒性试验、慢性毒性试验、致突变性试验和致癌性试验等。常用半数致死量LD50和半数致死浓度LC50两个参数评估急性毒性,而常用未观察到任何效应的最高剂量(no observed effect level,NOEL)和能观察到任何生物学效应的最低剂量(lowest observed effect level,LOEL)两个参数来评估慢性毒性。为全面评估SDHI类杀菌剂对生物体的毒性效应和潜在风险,本文主要针对其急性毒性进行分析,研究方法包括实验室模拟、田间观测和风险评估等,而关于SDHI类杀菌剂的生态毒性研究主要聚焦于动物,对其毒性的分级主要采用《化学农药环境安全评价试验准则》(GB/T 31270.17—2025)。
在大部分研究中SDHI类杀菌剂对多种水生生物表现出强烈的致死作用,其急性毒性普遍为中毒至剧毒,仅少部分为低毒。例如,SDHI类杀菌剂苯并烯氟菌唑、吡唑萘菌胺和氟唑环菌胺会导致斑马鱼Danio rerio胚胎形态发生变化,孵化失败和死亡,也会干扰胚胎发育、心脏发育、先天免疫等相关基因的表达,从而导致鱼体较短或心跳速率紊乱等;同时,慢性暴露于上述3种SDHI类杀菌剂会影响斑马鱼雌成鱼的氧化应激、内分泌和神经系统,进而影响其能量代谢和肠道组织形态,表明这3种SDHI类杀菌剂对斑马鱼的胚胎和成鱼均表现出急性毒性。暴露噻呋酰胺后,斑马鱼比肝重、比鳃重均显著升高,可能暗示噻呋酰胺抑制肝脏和鱼鳃中SDH及线粒体呼吸链复合物Ⅱ的活性,从而影响电子转移和能量供应,造成机体能量供应不足,进而影响斑马鱼的身体循环功能或对组织器官造成损伤,最终导致其不能正常孵化,甚至死亡。氯氟联苯吡菌胺和苯并烯氟菌唑对鱼类也有剧毒,LC50介于0.004~0.095 mg/L之间,会导致斑马鱼胚胎及其发育阶段的仔鱼形态异常,如出现心包囊水肿、卵黄囊水肿等症状,同时伴有小头畸形、脊索弯曲以及尾部弯曲等症状,其中苯并烯氟菌唑毒性最强。苯并烯氟菌唑不仅对鱼类有剧毒,而且对大型水溞Daphnia magna也有剧毒,LC50为0.085 mg/L,其中在暴露初期,受试生物会出现快速且不规则的运动模式,对环境变化的感应能力显著下降,肢体出现细微震颤,且逐渐沉降至培养容器底部;随着暴露时间的累积,上述症状愈发严重,中毒个体数量呈上升趋势;24 h后超过半数的个体死亡,且死亡个体的体色变为苍白。苯并烯氟菌唑对鱼类和大型溞均有剧毒,必须严格控制其浓度,以确保水生生物的健康和生态环境的平衡。啶酰菌胺对羊角月芽藻Selenastrum capricornutum、大型溞和斑马鱼的毒性分别为低毒、中毒和中毒。试验时上述杀菌剂均处于行政保护期,当不受行政保护后,这些杀菌剂的用量可能会大量增加,可能对水生生物的毒害更大。吡唑萘菌胺、氟唑菌酰胺和氟唑菌酰羟胺对鱼类有高毒,LC50介于0.121~0.545 mg/L之间。新杀菌剂茚吡菌胺对斑马鱼胚胎有强毒性,处理96 h后的LC50为0.063 mg/L,而0.050 mg/L浓度即可导致胚胎卵黄囊水肿、心包水肿和脊柱弯曲。氯氟联苯吡菌胺对水藻的EC50为0.097 mg/L,远低于0.300 mg/L的高毒标准,这意味着即使在极低浓度下氯氟联苯吡菌胺也可能对水藻有强毒性,进而影响水生生态系统的平衡。氟唑菌酰胺、氟唑菌酰羟胺和异丙氟吡菌胺对大型溞的EC50介于0.201~0.700 mg/L之间,为高毒,能抑制大型溞的活动。氟唑菌酰羟胺对鱼类和大型溞均为高毒。萎锈灵、麦锈灵、氟酰胺、吡噻菌胺、氟唑环菌胺、噻呋酰胺和啶酰菌胺均对鱼类有中毒,EC50介于1.560~6.400 mg/L之间。氟茚唑菌胺可导致蚯蚓Lumbricus terrestris组织损伤,并引起其发生氧化应激反应,其中1 mg/kg浓度即可诱发蚯蚓肠道空泡化与肌纤维松散,暴露28 d后蚯蚓体内超氧化物歧化酶、谷胱甘肽酶耗竭,活性氧激增,呈现典型的氧化应激-组织损伤级联反应。
总而言之,SDHI类杀菌剂对非靶标生物的毒性效应不容忽视。SDHI类杀菌剂的生态毒性效应广泛且复杂,其环境行为和潜在风险需进一步深入研究。目前关于SDHI类杀菌剂对多数非靶标害虫和哺乳动物的生态毒理研究较匮乏,在未来需深入研究。在实际应用中,为减轻SDHI类杀菌剂的生态毒性,必须严格控制其使用浓度,研发新剂型或开发更科学的实用技术,包括合理控制用药量和使用频率,选择低毒或环境友好型的杀菌剂品种,加强农药残留检测和监管等。此外,还需要加强生态风险评估和预警体系的建设,以便及时发现并应对潜在的生态风险。
4 SDHI类杀菌剂抗性问题及其应对策略
SDHI类杀菌剂因其高效、广谱的杀菌特性在全球农业生产中广泛应用。然而,随着使用频率的增加,病原菌对SDHI类杀菌剂的抗性问题逐渐凸显,已成为制约农业可持续发展的关键因素之一。抗性产生的原因主要归结于SDHI类杀菌剂的广泛使用和由此产生的选择性压力,这促使病原菌通过多种机制适应药剂环境,主要包括靶标酶突变(如SDH酶亚基突变)、非靶标酶突变(代谢抗性)以及其他机制(如生物膜形成、基因表达调控)。目前,多种病原菌已对SDHI类杀菌剂产生抗性,且抗性发展迅速,全球范围内均有报道。例如,引起植物灰霉病的主要病原菌灰葡萄孢菌已对SDHI类杀菌剂产生显著抗性,其抗性机制主要与SDH亚基的突变有关,如SDHB亚基中P225F、N230I、H272R/Y/L/V等位点突变以及SDHC亚基中H152R、H134R等位点突变,这显著降低了灰葡萄孢菌对SDHI类杀菌剂的敏感性。在西班牙、意大利以及中国的上海、江苏和重庆等地,灰葡萄孢菌对啶酰菌胺的抗性频率分别为7.60%、59.00%、29.74%、42.60%和92.25%。黄瓜靶斑病菌对氟唑菌酰羟胺和吡唑萘菌胺的敏感性显著下降,EC50分别为258.0 mg/L和23.77 mg/L,显著高于其敏感基线,其抗性下降的主要原因是,SDHB-I280V和SDHC-N75S菌株发生突变,突变后菌株在田间仍保持较高的生长活力。孙龙江通过室内药剂驯化获得了8株对苯并烯氟菌唑产生抗性的花生白绢病菌菌株,其抗性因子介于21.58~35.08之间,抗性程度为中等;此外,抗性菌株与亲本菌株或同一亲和群的菌株菌丝融合后对苯并烯氟菌唑的敏感性显著降低,从而增加了对大花生白绢病的防治难度。
未来还需深入研究抗性机制,利用基因组学、转录组学、蛋白质组学和代谢组学等多组学技术全面解析病原菌对SDHI类杀菌剂的抗性机制,探索新的抗性机制和靶点。此外,研发新型SDHI类杀菌剂也是应对抗性问题的重要途径。通过对SDHI类杀菌剂结构的优化提高其对病原菌的活性和选择性,降低抗性风险。探索新的靶点和作用机制,开发具有不同作用机制的新型杀菌剂,以满足农业生产的需求。在新型SDHI类杀菌剂的研发过程中,注重绿色化学理念的应用,减少对环境的污染和对生态系统的破坏,实现农业的可持续发展。
5 SDHI类杀菌剂的环境行为及分析方法
5.1 SDHI类杀菌剂的环境行为
SDHI类杀菌剂使用后,其可能在植物体内或土壤中残留。SDHI类杀菌剂在土壤中的行为主要受吸附与解吸、降解以及土壤质地、有机质含量和pH值等土壤物理和化学性质的影响。例如,宋明霞等研究发现土壤类型、湿度、氧气条件等因素均会影响吡唑萘菌胺的吸附,进而影响降解速率;在黑土、水稻土、潮土和红土4种土壤中,吡唑萘菌胺的降解速率由大到小依次为黑土、水稻土、潮土和红土;土壤湿度越大,吡唑萘菌胺降解得越快;厌氧条件下吡唑萘菌胺的降解比好氧条件下快。樊雨鑫等通过室内试验发现,土壤中有机质和酸性土壤条件能加速苯并烯氟菌唑的降解过程,因为有机质丰富的土壤为苯并烯氟菌唑提供了更多的吸附位点,而酸性条件土壤对苯并烯氟菌唑有吸附作用。吴文铸等通过试验发现,环氟菌胺的降解速率随着土壤pH的升高而加快,因为pH可以改变土壤中微生物的活性和农药的电荷状态,从而影响农药的降解。此外,土壤微生物和酶的活动也参与SDHI类杀菌剂的降解过程。例如,谢海燕研究表明,在不同pH的土壤中吡噻菌胺的降解半衰期受土壤微生物和酶活性的影响,其中,在酸性土壤中其降解速度最快,在近中性土壤次之,而在碱性土壤中降解速度最慢;在碱性土壤中降解最缓慢主要是因为在碱性条件下吡噻菌胺结构比较稳定,同时微生物的活性也受到一定限制。
杀菌剂对水体的污染方式有直接向水体施药、随雨水向水体中迁移、农药生产和企业加工时排放、大气残留农药随水进入水体等多种方式。SDHI类杀菌剂的残留可能导致农产品中农药残留超标,同时也可能通过植物生长和繁殖过程进入土壤和水体,进而影响人类健康。尽管其挥发性相对较低,但SDHI类杀菌剂仍可能通过地表径流和大气漂移进入水体,如在模拟暴雨径流中氟酰胺的浓度可达798μg/L。SDHI类杀菌剂还可能通过生物富集和生物放大作用在食物链中逐级积累。因此,需从多途径暴露、生物富集和生物放大效应等多方面全面评估SDHI类杀菌剂对环境的风险,并采取措施减少其对环境和人类健康的潜在危害。SDHI类杀菌剂在环境中的部分降解产物也会对环境造成危害。例如,向发椿研究发现氟唑菌酰胺在环境中主要通过水解和生物降解途径降解,其主要降解产物为M700F001和M700F002,这些降解产物的毒性相对较低,但在水生生态系统中仍可能有一定的生态毒性。此外,氟唑菌酰胺在土壤中的降解半衰期较长,其可能通过地表径流和土壤淋溶进入水体,在水生环境中SDHI类杀菌剂及其降解产物可能通过食物链传递对水生生物产生潜在影响。例如,某些SDHI类杀菌剂的降解产物在水生生物体内的生物富集因子较高,表明其有一定的生物富集能力。因此,在评估SDHI类杀菌剂的环境风险时,需要综合考虑其降解产物的种类、毒性以及在环境中的归趋,从而制订有效的污染控制策略。
土壤被污染后,SDHI类杀菌剂也会通过上述方式渗透进入水体,对水环境造成破坏,进而危害水生生物。不同种类SDHI类杀菌剂经一系列过程进入到水体后会对水生生物的机体功能造成不同的影响,甚至有致死效果。在使用SDHI类杀菌剂时应重视其在土壤中的降解处理和在水中的净化过程,尽量减少其对水环境与水生生物的为害。因此,了解SDHI类杀菌剂的环境行为有助于评估其对生态系统的潜在影响,从而确保其在农业应用中的可持续性和环保性。
5.2 SDHI类杀菌剂的分析方法
SDHI类杀菌剂的分析方法有液相色谱-串联质谱(liquid chromatography-tandem mass spectrom‐etry,LC-MS/MS)法、超高效液相色谱-四级杆/静电场轨道阱高分辨质谱(ultra-performance liquid chromatography-quadrupole-electrostatic field Orbitrap high-resolution mass spectrometry,UPLC-Q-Orbitrap HRMS)法、气相色谱(gas chromatography,GC)法、气相色谱-质谱(gas chromatography-mass spectrometry,GC-MS)法等;SDHI类杀菌剂的分析范围主要涵盖植物源、动物源、食用菌、环境等介质,其中对植物源的检测较多。例如,毕思远等将QuEChERS(quick,easy,cheap,effec-tive,rugged,safe)方法与GC-MS相结合来检测葡萄浆果中噻呋酰胺的残留。龚蕾等改进QuEChERS和净化方法,并将其与UPLC-MS/MS相结合测定果蔬中18种SDHI类杀菌剂的残留量,此方法操作简单,净化效果佳,可用于果蔬中新型SDHI类杀菌剂的快速检测。边海涛等采用LC-MS/MS法检测柑橘、土豆、大豆及水果加工产品等不同基质中上啶酰菌胺、氧化萎锈灵、灭锈胺、吡噻菌胺等近百种农药或其代谢物的残留。汤晨采用LC-MS/MS法测定水产品中吡唑萘菌胺、苯并烯氟菌唑、氟唑菌酰胺这3种SDHI类杀菌剂的残留量,该方法操作简单、灵敏度高和净化效果好。SDHI类杀菌剂在肉类中残留主要采用LC-MS/MS法、QuEChERS法与LC-MS/MS法结合和UPLC-Q-Orbitrap HRMS技术等方式进行检测;SDHI类杀菌剂在动物源中的残留量也采取QuECh-ERS法与LC-MS/MS法结合的方式检测。例如,张丽娟等用此方法快速测定猪肉、鸡肉、鸭肉、牛肉等肉类中8种新型SDHI类杀菌剂的残留量,检出限介于0.3~1.5μg/kg之间,回收率为80.5%~103.5%,相对标准偏差为1.3%~6.5%。胡均鹏等研究也表明此方法能简单、快速、高效地检测肉类食品中9种SDHI类杀菌剂的残留量。古飞燕等利用UPLC-Q-Orbitrap HRMS技术建立了一种高效筛选动物肌肉组织中16种新型SDHI类杀菌剂残留的方法,试验结果表明,该方法具有灵敏度高、准确性高、检出限低的优点,平均回收率介于67.1%~111.9%之间,相对标准偏差介于0.3%~12.0%,适用于动物肌肉组织中新型SDHI类杀菌剂残留的检测。李敏青等使用QuECh-ERS-LC-MS/MS法同时测定动物源性食品中21种SDHI类杀菌剂的残留量。GC法和GC-MS法还可以用于测定土壤、沉积物中SDHI类杀菌剂的残留。例如,段劲生等采用GC法对水稻田中呋噻酰胺的残留进行检测,其中,呋噻酰胺在土壤、植株、谷壳、糙米中的平均添加回收为73.6%~99.8%,最小检出量为5.0×10-12 g。许晓霞采用GC-MS测定土壤中甲呋酰胺、吡噻菌胺、麦锈灵、灭锈胺和呋吡菌胺5种SDHI类杀菌剂的残留量。在25种SDHI类杀菌剂中,关于异丙氟吡菌胺、茚吡菌胺等最新一代杀菌剂残留方法的报道较少见。
综上所述,常采用LC-MS/MS、GC、GC-MS等方法来检测SDHI类杀菌剂残留,其中QuEChERS法与LC-MS/MS法相结合在SDHI类杀菌剂中应用最广,可用于果蔬、肉类以及咖啡中SDHI类杀菌剂残留的检测。深入研究SDHI类杀菌剂的环境行为和开发高效的分析方法,对于科学评估其对生态系统和环境的潜在风险至关重要。
6 展望
SDHI类杀菌剂作为现代农业生产中具有高效杀菌活性的核心药剂之一,其在生态毒理、剂型创新、抗性机制、健康风险等方面面临关键挑战。首先,加强SDHI类杀菌剂环境行为归趋、代谢产物毒理研究,尤其是对土壤微生物、陆生无脊椎动物等非靶标生物的生态毒理学数据挖掘,建立多维生态风险评估体系。其次,加快推进SDHI类杀菌剂复配制剂与绿色剂型创新,提升靶标生物活性,通过科学复配延缓抗性演化,研发微胶囊、水乳剂等环境友好型剂型,降低农药残留环境负效应。此外,结合智能化现代农业技术构建精准施药体系,通过科学轮换用药、控制施用频率与间隔等措施实现生物活性与生态安全的协同提升,持续为现代农业高质量发展与绿色农药创新应用提供科学支撑。
