
摘自:《生物技术通报》
作者:金曼1 龚奕杭2 潘文波2 罗培润1 高伟2 李平东*1 唐晓艳*2,3(1.深圳市农产品质量安全检验检测中心 2.华南师范大学生命科学学院广东省植物发育生物工程重点实验室 3.深圳市作物分子设计育种研究院)
水稻作为全球尤其是亚洲地区最主要的粮食作物之一,其生产安全对保障世界粮食供应至关重要。然而,稻田杂草的危害严重威胁着水稻的产量和品质。
在抗除草剂水稻育种领域,国内外科学家采用化学诱变等传统技术,成功培育出多个非转基因抗除草剂的水稻品种。目前,以乙酰乳酸合成酶(ac⁃etolactate synthase, ALS)为作用靶标的抗除草剂水稻应用最为广泛,如巴斯夫的Clearfield®水稻以及深圳洁田模式生物科技有限公司的洁田稻等。然而,通过诱变获得抗除草剂水稻材料存在周期长、效率低等缺点。通过基因组编辑技术可以对水稻基因组进行精确定点编辑,获得目标位点突变的水稻材料,从而创制具有除草剂抗性的水稻种质。
基因组编辑技术的核心原理是利用核酸酶在基因组的特定目标位点,精准诱导 DNA 双链断裂(double-strand break,DSB)。当 DNA 双链出现断裂后,细胞会自主启动修复机制,通过同源定向修复(homologous-directed repair,HDR)或非同源末端连接(non-homologous end joining,NHEJ)两种途径对断裂处进行修复,在修复的过程中实现基因序列的改变。在该技术发展早期,所使用的序列特异性核酸酶包括归巢核酸内切酶(meganuclease)、锌指核酸酶(zinc-finger nucleases,ZFNs)、转录激活样效应因子核酸酶(transcription activator-like effector nucleases,TALENs)等。尽管它们具备编辑植物基因组的能力,但是由于组装费时、成本高和基因编辑效率低等因素限制了其应用范围。近年来兴起的CRISPR/Cas(clustered regulatory interspaced short pal⁃indromic repeat/CRISPR-associated protein)系统编辑效率高、设计和操作简单、成本低、适用范围广,已成为当前主导的基因编辑技术,并成功运用于多种作物基因组的定向编辑。
基因编辑在创制抗除草剂水稻中的应用
抗乙酰乳酸合成酶(ALS)抑制剂类水稻
乙酰乳酸合成酶是植物中支链氨基酸合成过程中的一种关键酶。ALS 抑制剂类除草剂通过抑制 ALS 的活性使其无法正常发挥功能,进而阻止植物体内支链氨基酸的合成,导致植物的蛋白质合成、细胞分裂过程受到干扰,生长发育陷入异常,最终导致植物死亡。截至目前,全球范围内已发现 176 种杂草对ALS抑制型除草剂产生抗性,产生抗性的原因主要是由于ALS基因编码区氨基酸的突变。以拟南芥ALS氨基酸序列为参照标准,已在抗性杂草中鉴定出 8 个氨基酸突变位点,分别为 Ala122、Pro197、Ala205、Asp376、Arg377、Trp574、Ser653和 Gly654。这些突变位点会使杂草对不同类型的ALS 抑制剂类除草剂产生不同程度的抗性,包括咪唑啉酮类(imidazolinones,IMI)、磺酰脲类(sulfo⁃nylureas,SU)、三唑并嘧啶类(triazolopyrimidines,TP)、嘧啶硫苯甲酸类(pyrimidinylthiobenzoates,PTB)以及磺酰氨基羰基三唑啉酮类(sulfonylamino⁃carbonyltriazolinones,SCT)等。
研究人员通过靶向编辑OsALS基因的保守位点,获得了丰富多样的突变形式(表1)。Wang等通过 NHEJ 修复途径对 OsALS 基因进行靶向编辑,使OsALS基因第1882位的碱基发生G到T的颠换,导致628位Gly 突变为 Trp,从而使编辑后的水稻对咪唑烟酸(imazapic)以及咪唑乙烟酸(imazethapyr)产生很高的抗性。还有研究人员通过 HDR 途径对OsALS进行基因编辑,一种是以含有Trp548Leu突变或含有 Trp548Leu 和 Ser627Ile 双突变的 DNA 为模板进行基因编辑;另一种是以编码Trp548Leu和Ser627Ile双突变的RNA为模板进行基因编辑, 通过两种方式均获得了对双草醚(bispyribac)具抗性的水稻材料。
表1 利用基因编辑获得的抗乙酰乳酸合成酶(ALS)抑制剂类水稻
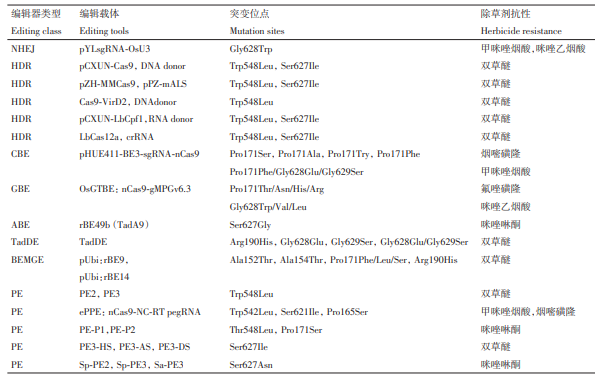
通过碱基编辑,Zhang等利用CBE,针对水稻OsALS 基因的Pro171和Gly628位点进行编辑,获得Pro171Ser/Ala/Try/Phe单突及Pro171Phe/Gly628Glu/Gly629Ser 三突类型。Tian 等开发的 OsGTBE 工 具通过独特的 G 切除机制,实现了水稻中高效的 G到 T 碱基编辑,并在上述两个位点产生 Pro171Thr/Asn/His/Arg 和 Gly628Trp/Val/Leu 新突变类型,对咪唑乙烟酸和氟唑磺隆产生抗性。有研究人员利用ABE 对 OsALS 基因进行定向突变,成功获得了Arg190His、Ser627Gly、Gly628Glu、Gly629Ser和Gly628Glu/Gly629Ser等抗除草剂突变形式。此外,Kuang 等提出了一种基于碱基编辑的基因进化方法BEMGE,该方法利用CBE和ABE以及覆盖目标基因全长编码区的 sgRNA文库,实现OsALS的人工进化,产生 4 种不同类型的氨基酸突变类型Pro171Phe/Leu/Ser、Arg190His,赋予水稻对双草醚不同程度的耐受性。
利用引导编辑系统对 OsALS 进行编辑,通过对逆转录模板进行碱基替换,研究人员成功在水稻中引入Trp542Leu、Ser621Ile、Pro165Ser、Pro171Ser、Ser627Ile以及Ser627Asn等突变位点,获得了对咪唑啉酮、双草醚、甲咪唑烟酸、烟嘧磺隆等不同抗性的水稻材料。
抗乙酰辅酶A羧化酶(ACCase)抑制剂类水稻
乙酰辅酶A羧化酶(ACCase)在生物体的物质代谢过程中承担着关键催化功能,其核心作用是催化乙酰辅酶A发生羧化反应,生成丙二酰辅酶A。丙二酰辅酶A作为重要的中间代谢产物,为脂肪酸以及许多次生代谢产物的合成提供底物。因此,ACCase也是脂肪酸合成途径的限速酶。ACCase抑制剂类除草剂通过抑制ACCase来干扰脂肪酸的合成,进而阻碍膜系统中脂质成分的生成,造成细胞结构的破坏,最终导致植物死亡。ACCase抑制剂类除草剂包括芳氧苯氧丙酸酯类(aryloxy phenoxy propionate, APPs),环己二酮类(cyclohexanediones, CHDs),以及苯基吡唑啉类(phenylpyrazolin, DEN) 三类。质体 ACCase的羧基转移酶(carboxyl transferase, CT)结构域是这些除草剂的作用靶标区域。对抗性杂草的研究发现,在 CT 结构域中存在Ile1781Leu、Trp1999Ser/Cys、Trp2027Cys、Ile2041Asn、 Asp2078Gly、Cys2088Arg、Gly2096Ala氨基酸位点的突变,从而赋予了植物对不同 ACCase 抑制剂类除草剂的抗性,其中Ile1781Leu位点突变在植物中应用最广泛。
利用碱基编辑器对OsACCase基因的上述保守位点进行编辑,获得了众多的突变形式(表2)。研究人员利用ABE对OsACCase基因进行编辑,在靶区域检测到 A 到 G 替换,效率 15.8%-59.1%,且未检测到碱基插入、缺失或非预期编辑。通过氟吡甲禾灵筛选获得了Cys2186Arg突变的抗性水稻植株。此外,研究人员利用新型的饱和靶向内源诱变编辑器(STEMEs)、双腺嘌呤和胞嘧啶编辑器(ACEs)以及多重正交碱基编辑器(MoBE)等,通过构建sgRNA 文库,对水稻中的 OsACCase 进行定向进化,经除草剂筛选获得Ile1879Val、Pro1927Phe、Trp2125Cys/Ser/Leu/Gln、Ile2139Asn、Cys2186Agr/His等位点突变类型的抗性水稻材料。这些位点突变后,通过改变OsACCase蛋白与除草剂的结合构象,赋予水稻对氟吡甲禾灵的抗性。
表2 利用基因编辑获得的抗乙酰辅酶A羧化酶(ACCase)抑制剂类水稻
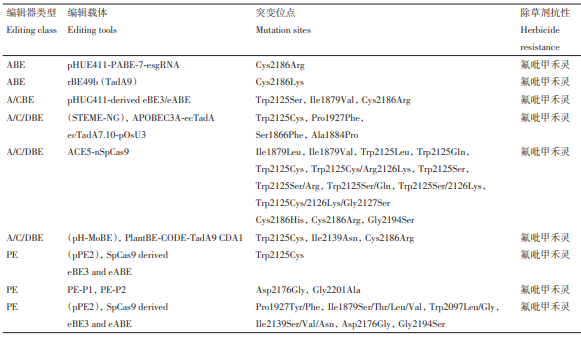
Xu 等率先验证了植物 PE 系统的可行性,通过 pPE 对 OsACCase 基因进行编辑,成功引入Trp2125Cys突变,获得了抗氟吡甲禾灵的水稻植株。该研究团队开发的引导编辑文库介导的饱和突变方法(prime-editing-library-mediated saturation mutagen⁃esis,PLSM),针对OsACCase上6个与除草剂敏感性相关的保守位点(Ile1879、Pro1927、Trp2097、Ile2139、Asp2176、Gly2194),设计含随机密码子的pegRNA 文库,构建 pPE2 载体质粒库并转化水稻愈伤组织,筛选获得抗氟吡甲禾灵的水稻材料。此外,利用 PE-P1 和 PE-P2 两个引导编辑系统,对OsACCase 的 Asp2176 和 Gly2201 两个位点实现抗性位点的精准突变,创制抗氟吡甲禾灵的水稻。
抗草甘膦水稻
草甘膦(glyphosate)是一种具有稳定 C-P 键的磷酸盐化合物,是以植物叶绿体中5-烯醇式丙酮酰莽草酸-3-磷酸合成酶(5-enolpyruvoylshikimate-3-phosphate synthase, EPSPS)为靶标的除草剂。作为叶绿体中的关键酶,EPSPS 在催化磷酸烯醇丙酮酸 (phosphoenolpyruvate, PEP)和磷酸莽草酸(shi⁃kimate-3-phosphate, S3P)生成 5-烯醇式丙酮酰莽草酸-3-磷酸(5-enolpyruvyl-shikimate-3-phosphate, EPSP)的反应中起到重要作用。该催化反应为植物合成芳香族氨基酸以及激素、类黄酮等次生代谢物质的关键步骤。喷施的草甘膦与PEP竞争结合EPSPS活性位点,抑制酶的催化功能,进而阻断莽草酸途径:一方面蛋白质生物合成所必需的芳香族氨基酸合成严重受阻,莽草酸过量积累;另一方面干扰植物生长所需的激素以及次生代谢物质的合成,从而引起生长代谢紊乱,导致植物死亡。
在牛筋草(Eleusine indica)、黑麦草(Lolium rigidum)、多花黑麦草(Lolium multiflorum)、两耳草(Digitaria insularis)、糙果苋(Amaranthus tuber⁃culatus)和稗草(Echinochloa colona)等杂草中发现,EPSPS蛋白酶上102位Thr突变为Ile,103位Ala突变为Val,以及106位Pro突变为Ser、Ala、Thr或者 Leu 后可以使该酶对草甘膦产生抗性,导致喷施大田剂量的草甘膦后杂草仍能生存。利用这些抗性位点,研究者们通过基因编辑手段将水稻的 EPSPS 位点进行改造,从而获得具有草甘膦抗性的水稻材料(表3)。
表3 利用基因编辑获得的抗草甘膦水稻
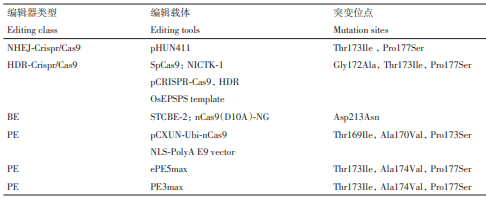
Li 等开发了高效的内含子介导的位点特异性基因替换和插入方法。该方法借助 CRISPR-Cas9系统,通过NHEJ修复途径使得基因产生突变。利用靶向相邻内含子的 sgRNA 对及同源供体 DNA,以2.0%频率实现OsEPSPS基因片段替换,而靶向单个内含子的sgRNA及同源供体DNA介导的基因插入效率为2.2%。实验结果表明,携带了含有预期替换的OsEPSPS 基因的水稻植株具有抗草甘膦的特性。Sony 等利用 CRISPR-Cas9 技术,通过构建含有水稻密码子优化的SpCas9的双sgRNA的载体,与含有突变位点的OsEPSPS-HDR模板共转入水稻中,成功对 Gly172Ala、Thr173Ile 和 Pro177Ser 位点进行替换,获得含有GATIPS突变的水稻材料。该水稻材料表现出对草甘膦的抗性,同时增加芳香族氨基酸含量并提高稻谷产量。
Zhang等将高效的胞嘧啶脱氨酶(evoFERNY)和腺嘌呤脱氨酶(TadA8e)与 nCas9-NG(可识别NG型PAM序列)融合,并在C端添加两个UGI,构建了STCBE-2系统。针对OsEPSPS基因设计了35条sgRNA 并分为 11 组,通过该系统介导近饱和突变,经草甘膦和潮霉素双重筛选,获得含 Asp213Asn 新突变形式。该突变改变了草甘膦结合域的结构,降低了酶与草甘膦的结合能力,其转基因纯合株系在4倍推荐剂量的草甘膦处理下仍能正常生长,而野生型死亡。
研究发现,在 OsEPSPS 中引入Thr169Ile、Ala170Val 和 Pro173Ser(即TIAVPS)三重氨基酸突变,可能会使水稻对草甘膦产生更高水平的抗性。Li 等利用引导编辑对 EPSPS 基因进行TIAVPS位点突变,获得了45株独立的转基因植株。在这些植株中,筛选到一个杂合株系,其中一个等位基因精确编辑为TIAVPS突变类型,而另一个等位基因在 pegRNA 和 sgRNA 靶位点之间有一个 10 个碱 基对的插入,编辑效率为 2.22%。ePE5max 在诱导OsEPSPS生成纯合TIAVPS突变方面表现更优,纯合突变效率达 6.1%,并成功赋予水稻对草甘膦的抗性。Gupta等建立了多个epegRNA & ngRNA表达框的载体系统,可对水稻进行高效编辑,产生OsEPSPS TIAVPS突变类型。
抗草铵膦水稻
草铵膦是一种非选择性广谱除草剂,特异性作用于植物谷氨酰胺合成酶(glutamine synthetase, GS)。在植物氮代谢过程中,GS的核心作用是催化谷氨酸和氨转化为谷氨酰胺,是该催化反应中的关键酶。作为谷氨酸的类似物,草铵膦会与GS的天然底物竞争结合位点,从而抑制植物的氮同化作用,导致氨在植物体内过量积累,进而引发光合活性下降、叶绿体结构破坏、乙醛酸降解,并最终导致植物死亡。
抗草铵膦杂草的出现相对较缓。截至目前,仅发现1种牛筋草和3种黑麦草对草铵膦表现出一定程度的抗性。当意大利黑麦草 GS 的 171 位 Asp 被Asn 取代后,对草铵膦产生一定的抗性。进一步的研究表明,该氨基酸位点突变后GS对草铵膦敏感性下降,其原因之一可能是氨基酸极性发生改变。对紫花苜蓿GS的研究显示,当该蛋白207位氨基酸为 Gly、245 位为除 Gly 外的任意氨基酸、332 位为Arg或Lys时,会表现出一定程度的草铵膦抗性。
Ma 等利用 CRISPR/Sc++系统介导的 rBE73b对 OsGS1 基因进行编辑,在 40 个独立株系中,有 9个在目标位点发生A至G转换,导致131位氨基酸由 Glu 转换为 Gly。Yan 等利用 rBE49b 对 OsGS1 基因的His249密码子 进行靶向编辑,并在靶区域发生 A 到 G 的编辑。Ren 等利用 BEMGE 系统,设计 102 个 sgRNA,构建碱基编辑文库,并通过农杆菌导入粳稻品种Kitaake,经筛选基因分型确定获得3个赋予草铵膦耐受性的新型OsGS1等位基因,即OsGS1-Ala63Val/Pro64Ser、OsGS1-+65Ala/66Phe 和OsGS1-Ser59Gly/Thr60Ala。研究人员采用重离子束处理、乙基甲基磺酸诱变等技术,结合草铵膦筛选,获得一系列抗草铵膦水稻突变体(表4)。其中,OsARF18(LOC_Os06g47150,)突变后的水稻材料对草铵膦具有一定的抗性。
表4 利用基因编辑获得的抗其他类型除草剂水稻
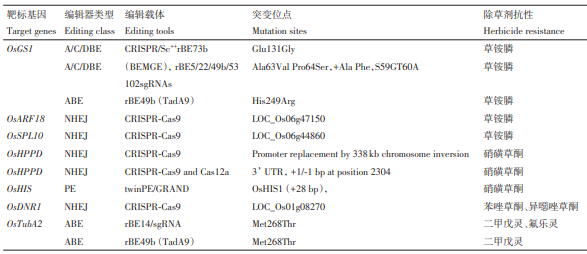
研究人员利用 CRISPR-Cas9 对水稻OsARF18进行敲除,获得了具有草铵膦抗性且对产量无负面影响的水稻材料。研究表明,草铵膦可诱导ARF18基因的表达,进而抑制参与氨和活性氧(reactive oxygen species, ROS)清除的下游基因的表达,导致体内氨和ROS过量积累引发植物死亡。而ARF18基因突变后,下游相关基因正常发挥功能,及时清除积累的氨和ROS,以抵御草铵膦带来的损伤。此外,在突变体库中分离获得的gar6-2突变体,在LOC_Os06g44860(OsSPL10)第685位碱基由C突变为T,使得水稻获得草铵膦抗性。OsSPL10作为草铵膦抗性的负调控因子,通过调节水稻GS活性发挥作用。利用CRISPR-Cas9基因编辑技术构建的水稻OsSPL10敲除系在草铵膦处理下存活率达100%,而野生型全部死亡,为作物草铵膦抗性改良提供了潜在候选基因。
抗HPPD抑制剂类水稻
对羟基苯基丙酮酸双加氧酶(4-hydroxyphenyl⁃pyruvate dioxygenase, HPPD)的核心功能是催化羟基苯丙酮酸(4-hydroxyphenylpyruvate, HPPA)转化为尿黑酸(homogentisate, HGA),而尿黑酸是植物体内质体醌和生育酚生物合成的前体物质。HPPD抑制剂类除草剂的作用机制是通过与酶活性中心的Fe²⁺发生螯合反应,阻碍HPPD与底物HPPA的结合,从而以竞争性的方式抑制 HPPD 的功能。在植物的生长代谢中,质体醌和生育酚是类胡萝卜素生物合成的前体,而类胡萝卜素在光合作用中具有重要作用。当HPPD活性被抑制后,光合电子传递被阻断,最终导致植物白化死亡。目前,长芒苋是主要对 HPPD 抑制剂类除草剂产生抗性的杂草。研究表明,抗性与敏感长芒苋的 HPPD 序列完全一致,但抗性植株中 HPPD 的表达量升高,这可能是其对HPPD抑制剂类除草剂抗性增强的原因。
通过 CRISPR-Cas9 对功能基因启动子或者非编码区(untranslated regions, UTR)序列进行编辑以改变目的基因的表达量,是近年来作物性状改良的重要手段之一(表4)。Lu 等通过 CRISPR-Cas9 技术将 OsHPPD 的启动子更换为 OsUbiquitin2,成功将OsHPPD 的表达量提高几十倍,并显著提高了水稻的硝磺草酮抗性。Wu 等在 OsHPPD 3' UTR 中选择了 10 个靶位点,构建了含有 10 个 sgRNAs 的CRISPR-Cas9和CRISPR-Cas12a载体。通过转化扬粳3012,筛选获得了在OsHPPD 3' UTR的第2304位分别有1个碱基缺失、1个碱基插入的两个抗性材料,从而创造出具有硝磺草酮抗性且不影响正常生长的水稻材料。
通过对非靶标基因HIS1以及OsDNR1进行编辑,也可获得抗HPPD抑制剂类除草剂水稻(表4)。Maeda等在水稻中发现HIS1基因,其编码的Fe²⁺-氧戊二酸的加氧酶,通过催化β-三酮类除草剂的羟基化解除毒性。含有HIS1的水稻对苯并双环酮(benzobi⁃cyclon, BBC)及其他 β-三酮类除草剂具有一定的抗性。而对 HPPD 抑制剂类除草剂敏感的水稻品种,在其HIS1编码区存在28 bp的缺失。通过引导编辑,在HIS1基因缺失的MingHui86水稻中插入28 bp碱基序列,筛选获得13个(效率为15.5%)对硝磺草酮具有抗性的水稻材料。此外,研究发现,水稻dnr1 突变体对 HPPD 抑制剂类除草剂具有抗性,敲除OsDNR1可提高水稻体内HPPA水平,竞争性减少除草剂与HPPD的结合,促进HGA合成,从而对苯唑草酮、异噁唑草酮等多种 HPPD 抑制剂类除草剂均表现出显著抗性。
抗二硝基苯胺类除草剂水稻
二硝基苯胺类除草剂广泛应用于大豆和棉花的除草,不仅能防除一年生禾本科杂草,还可控制部分一年生阔叶杂草。这类除草剂包括氟乐灵、二甲戊灵和乙丁氟乐灵等,通过与微管蛋白结合、干扰微管的聚合,从而抑制细胞分裂与伸长,最终导致植物死亡。相比其他除草剂,杂草对二硝基苯胺类除草剂产生抗性的概率较低,已发现微管蛋白Thr239Ile 和 Met268Thr 突变的牛筋草(Eleusine in⁃dica)、Leu136Phe 和 Thr239Ile 突变的狗尾草(Setaria viridis)、Val202Phe、Leu125Met、Leu136Phe 突变的看麦娘(Alopecurus aequalis)以及携带 Val202Phe、Thr239Ile、Arg243Met/Lys 突变黑麦草(Lolium perenne)等少数杂草对其产生抗性。
Yan 等,Liu 等选择水稻中 α -微管蛋白同源基因 OsTubA2 作为靶标,利用水稻腺嘌呤碱基编辑器 rBE14、rBE49b 对靶位点进行精准碱基编 辑,在水稻内源 OsTubA2 基因中引入 Met268Thr 突变,成功获得对二硝基苯胺类除草剂(氟乐灵、二甲戊灵)具有抗性且对生长无影响的新型水稻材料(表4)。
未来研究方向
基因编辑器的优化改良与编辑效能提升
目前 CRISPR/Cas9 系统虽已广泛应用于科研与育种领域,但在提高编辑效率、拓宽编辑范围、降低脱靶效应等关键技术层面仍有较大的优化空间。当前,新型基因组编辑技术的持续突破正不断拓展编辑能力的边界,除传统的基因敲除、点突变等基础操作外,新开发的碱基编辑工具通过扩大同一靶位点引入突变的多样性,显著提升了定向编辑的灵活性。引导编辑系统更进一步突破技术限制,允许将靶标氨基酸替换为所有可能的氨基酸变体,为系统筛选具有优异表型的等位基因提供了强大工具。在高效创制突变体方面,生殖细胞特异性基因组编辑系统通过定向富集突变细胞群,可快速生成数百万级别的突变体库,大幅提高了创制稀有高价值突变的概率。MutaT7 系统通过持续向靶基因引入突变,驱动基因实现迭代进化,直至获得符合育种需求的理想突变体。此外,通过精准创制基因组结构变异(如染色体倒位、片段重复 等),将高表达基因的启动子元件与目标基因精准连接,可实现目标基因的高效上调表达,为基因编辑技术在作物育种中的应用开辟了新路径。这些技术创新不仅丰富了编辑手段,更推动基因编辑从单一功能操作向系统化进化设计升级。
新型功能基因的挖掘与解析
在杂草抗性演化加剧和除草剂类型日益丰富的背景下,培育兼具广谱除草剂抗性与优良农艺性状的水稻品种已成为育种研究的核心方向。通过基因功能解析发现,部分关键基因的变异或调控可实现″一基因多抗″的育种价值。如OsALS蛋白Trp548Met位点突变后,对咪唑啉酮类、磺酰脲类、三唑并嘧啶类、嘧啶硫苯甲酸类以及磺酰氨基羰基三唑啉酮类除草剂均表现出强抗性,为广谱抗除草剂育种提供重要靶点;OsDNR1敲除株系对吡唑酮类、三酮类、异噁唑类等多种HPPD抑制剂均表现出强抗性,为培育″一基因多抗″品种奠定了基础。此外,敲 除OsDNR1基因后不仅能赋予水稻对HPPD抑制类除草剂的抗性,还能提高氮利用效率。这种″抗性—养分高效″的协同效应,为培育多优性状聚合的水稻品种开辟了新路径。
在作物育种实践中,产量、品质、抗性等关键性状间常存在遗传拮抗效应,深入挖掘此类″抗性—农艺性状双赢″的基因资源,对突破传统育种瓶颈具有重要意义。例如高抗性基因的导入可能伴随生长发育迟滞或产量下降,导致多性状协同改良面临″顾此失彼″的困境。为破解这一难题,需强化水稻功能基因组学的系统性研究:通过高通量基因编辑筛选、蛋白互作网络解析等技 术手段,精准识别具有多重调控功能的关键基因,阐明其在除草剂抗性通路与农艺性状形成网络中的分子机制。通过分子设计育种技术将优异等位基因精准聚合,实现除草剂广谱抗性与产量、品质、养分效率等农艺性状的协同优化。这种以功能基因为核心的育种策略,不仅能打破传统性状改良中的拮抗限制,更能为培育″少投入、多产出、环境友好″的可持续水稻品种提供科学支撑,推动水稻育种从单一性状改良向多维度综合提升跨越。
原文链接:https://doi.org/10.13560/j.cnki.biotech.bull.1985.2025-1321

